北极植物化石名录
- 1. 该名录是什么?
- 2. 该名录的一些专业词汇
-
- 2.1 taphoflora
- 2.2 florule
- 2.3 form (morphotype) identifier
- 2.4 NASR
- 2.5 AKSR
- 2.6 phytostratigraphy
- 2.7 floral phytostratigraphic phase
- 2.8 stratoflora
- 2.9 phytostratigraphic horizon
- 2.10 Cretaceous Arctic forests
- 2.11 formal names
- 2.12 informal names
- 2.13 reticulate evolution
- 2.14 specimen identifiers
- 2.15 pectinal veins
- 2.16 Arctic region evolve during the Cretaceous
- 3. 该名录的化石图片
- 4. 该名录的分类系统
1. 该名录是什么?
这本互动式名录首次汇集了阿拉斯加和俄罗斯东北部数以千计的植物化石插图,展示了古老的北极森林和北极失落的世界。这些化石的年代可以追溯到白垩纪和古新世,属于全球气候比现今更温暖的时期。它们为我们理解北极地区过去和未来可能的地质和生物进化提供了独特的档案。
你可以通过交互式地图获取这些化石的图片和发现地点的信息。此外,你也可以通过查找分类名称表(如果已知)、访问博物馆藏品或浏览相关化石的收藏家和研究人员的信息来浏览该网站。
值得一提的是,本网站由中国科学院植物研究所慷慨托管。
该名录网站地址由此进入。
2. 该名录的一些专业词汇
2.1 taphoflora
植物化石群(或区系群),指从一个或多个地层中埋藏的植物遗骸中提取的所有植物化石,这些植物化石是特定时间跨度内特定和有限区域内植被的特征。
2.2 florule
在阿拉斯加,Smiley使用 florule 一词来指特定地点的单个植物化石群。不清楚这是否包括来自几种不同沉积类型的几个埋藏层的组合,还是仅指一个埋藏层。根据其使用情况,它很可能是指从一个地理位置采集的化石,因此可能包括从几个不同沉积层和埋藏层中采集的材料。
2.3 form (morphotype) identifier
当化石尚未被正式鉴定时,会使用独特的数字形式标识符。这些数字前缀有字母字符,大致表示叶片结构的主要特征。
2.4 NASR
NASR 是北阿拉斯加区系亚区域的缩写。
2.5 AKSR
AKSR 是阿纳德尔-科里亚克区系亚区域的首字母缩写。
2.6 phytostratigraphy
植物地层学(Phytostratigraphy)是根据区系演化的时期划分来识别区域和次区域植物地层的方法。含有植物区系发展过程中一个区域或次区域阶段的taphoflora的沉积物代表一个地层。
2.7 floral phytostratigraphic phase
区域或次区域区系演化的一个阶段是由一个或多个特征基本相似的taphoflora组成的。一个阶段的所有植物群都有一系列共同特征(类群组合、植物群之间的定性和定量关系)。除了这些共性特征外,每一个阶段的taphoflora都有其自身的特殊性,反映了地理、生态和年龄上的独特性。
2.8 stratoflora
stratoflora “一词被定义为”…来自有限但相当大的区域内同时沉积的植物遗骸的一般分类组成,该区域经历了其自身的地质发展和有机世界的演化历史"。
2.9 phytostratigraphic horizon
植物地层学层位是由特定植物化石内容所特征化的所有同生层、它们的部分和辅助岩性地层亚分区组成。
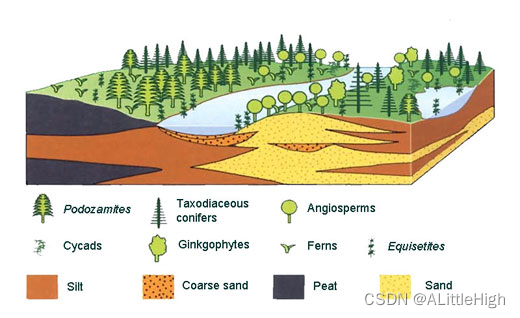
2.10 Cretaceous Arctic forests
在地球历史的过去2亿多年中,高纬度北极环境以森林而非冰川为主。对这些古代北极森林的组成、结构和动态有最详细了解的来源是在阿拉斯加和俄罗斯东北部发现的丰富植物化石。本目录提供了对这个已灭绝生态系统的自然档案的介绍。
开花植物(被子植物)首次出现在阿拉斯加和俄罗斯东北部的北极地区,出现在晚白垩世中晚期。它们加入了以落叶针叶树为主导的森林,次要的有银杏类植物和苏铁类植物。蕨类植物和石松类植物形成了地面植被覆盖层。这种类型的植被以库克波鲁克植物群为特征,具体描述可参考森林动态页面。
最早的北极被子植物群落占据了河流边缘的干扰场所,但很快在中等演替群落中多样化,在以针叶树为主导的森林中形成了林下植物群落,它们一直延续到古新世。下面是一个参考示意重建的塞诺曼期森林模型。
2.11 formal names
当标本被鉴定为先前描述的形态时,将使用正式的林奈二名法。林奈二名法由两部分组成,正如"二名法"一词所示:属名和种加词(例如Metasequoia occidentalis)。
2.12 informal names
形态类型标识符或“名称”通常在将叶子归属到已知属种困难时使用。它们是非正式但有用的方式,用于指代叶子类型,并基于一个系统,能够立即传达关于叶子形态的一些信息。
2.13 reticulate evolution
通常,进化被认为具有分支模式(“生命之树”),但在一类重要生物群体(例如开花植物)的早期进化过程中,频繁的杂交导致网状模式,而不是通常的分支模式。
2.14 specimen identifiers
这些差异取决于材料的来源。
2.15 pectinal veins
梳状脉是从叶片的中脉(或其他梳状脉)分离出来并在中央最多支持次级分支的侧脉(或侧脉)。
2.16 Arctic region evolve during the Cretaceous
在晚白垩世的早期阶段,板块运动消除了与太平洋的深水连接,而北冰洋在整个晚白垩世以及早新生代的大部分时间内相对封闭和与世界其他海洋隔离开来。
3. 该名录的化石图片
这是阿拉斯加和俄罗斯东北部白垩纪和古新世岩石中发现的标本插图的入口。并非每幅插图都附有相同水平的地点、年代和传统拉丁文名称数据。有些只有形态(形式)代码作为标识符。这是因为原始标本中缺少正式名称,或者材料尚待研究和描述。尽管如此,这里还是收录了这些标本,以尽可能全面地展示在北极地区发现的植物化石的多样性。
这里提供了关于图片页面布局、命名和形式标识符的指南。
图片按年代、分类名称、收藏和收藏者分组。要访问按地点排列的图片,请使用地图页面。
3.1 Age(年代)
这里的图片是根据化石所在岩石单元的年龄进行分组的。有些岩石单位的地层(年龄)范围有限,而其他岩石单位的年龄范围则不太明确。因此,不同年龄段的化石有不同大小的化石段。
- Paleocene (Selandian)
- Paleocene
- Maastrichtian-Paleocene
- Maastrichtian
- Campanian-Maastrichtian
- Early Campanian
- Santonian
- Santonian-Campanian
- Coniacian
- Turonian-Coniacian
- Turonian
- Cenomanian-Turonian
- Middle Cenomanian
- Cenomanian
- Late Albian-Cenomanian
- Late Albian
- Middle Albian-Cenomanian
- Middle Albian
- Early- Middle Albian
- Early Albian
- Unknown
按照不同的年龄段选择查阅对应的化石图片,如果结果太多,会按照字母表分段进行分页,而不是进行阿拉伯数字分页。每条结果会显示出化石产出地点、产出地层、年龄、分类、作者等信息,详细信息和图片在Link中。
3.2 Taxonomic Names
这些是按分类群组排列的图片快速链接。
- Angiosperms
- Bryophytes
- Conifers
- Cycadophytes
- Ferns
- Ginkgophytes
- Indeterminate
- Leptostrobaleans
- Lycophytes
- Caytoniales
- Sphenophytes
- Fauna
3.3 Collections
本目录中的化石收藏在美国和俄罗斯的多个机构中。这些不同收藏馆的图片可在此查阅。FGSM - 马加丹联邦地质调查博物馆,GINRAS - 莫斯科俄罗斯科学院地质研究所,MFGS - 马加丹联邦地质调查局,NEISRI - 马加丹东北综合科学研究所,USNM - 华盛顿特区史密森学会美国国家博物馆。
- FGSM
- GINRAS
- GINRAS 3384
- GINRAS 3385
- GINRAS 3390
- GINRAS 3823
- GINRAS 4843
- GINRAS 4894
- MFGS
- NEISRI-PF1
- NEISRI-PF3
- NEISRI-PF4
- USNM Angiosperms A-D
- USNM Angiosperms E-I
- USNM Angiosperms J-P
- USNM Angiosperms Q-Z
- USNM Algae?
- USNM Bryophytes
- USNM Conifers A-M
- USNM Conifers N-Z
- USNM Cycadophytes
- USNM Ferns A-C
- USNM Ferns D-Z
- USNM Ginkgophytes
- USNM Indet.
- USNM Leptostrobaleans
- USNM Lycopods
- USNM Pteridosperms
- USNM Sphenophytes
3.4 Collector
本目录中的图像资料由众多野外地质学家和古植物学家收集,代表了他们在偏远地区多年的野外工作时间。这里的图片按照收集者排列。
- Ahlberg
- Case
- Detterman
- Devyatilova
- Herman (Amaam, Arman R.)
- Herman (Grebenka GINRAS)
- Herman (Grebenka NEISRI)
- Herman (Sagavanirktok River)
- Hollick
- Lebedev, E.L.
- Lewis
- Moiseeva
- Parrish
- Pergament
- Philippova
- Raikevitch
- Rees
- Samylina
- Schorn
- Shchepetov
- Smiley (Corwin Bluffs)
- Smiley (Kuk River)
- Smiley (Kukpowruk River)
- Smiley (Lower Colville)
- Smiley (Pitmegea River)
- Smiley (Unknown, Chandler River)
- Smiley (Upper Colville)
- Spicer, A.J.
- Spicer, R.A. (Chignik, Grebenka)
- Spicer, R.A. (Ikpikpuk)
- Spicer, R.A. (Sagavanirktok River)
- Spicer, R.A. (Upper Colville)
- Terechova
- Union Oil
- Wolfe
- Zhuravlev
4. 该名录的分类系统
植物化石的命名既复杂又混乱。说它复杂是因为植物从未作为一个整体保存下来。叶片、花粉、花朵和种子都是在母体植物的整个生命过程中脱落的,而这些独立的部分会被单独制成化石。因此,古植物学家无法获得鉴定完整植物所需的全套特征。此外,特征的保存也并非完美无缺。在某些情况下,细胞的细节可能会保存下来,而在另一些情况下,整个器官(如叶片形状)的轮廓可能仅存于世。这些因素给植物化石的鉴定和正确命名带来了很大的不确定性。
多年来,这种不确定性造成了相当大的混乱。旧文献的作者倾向于将保存下来的任何特征与活体植物部分进行比较,这是可以理解的。当时几乎没有统一的规程,"图片匹配 "的文化逐渐形成,以至于活体(现存)属的名称,有时甚至是种的名称,都是根据最微弱的相似性应用到化石遗骸上的。这反过来又导致人们认为许多属和种都非常古老,而根据我们现在对进化过程的了解,以及随着时间的推移地理和气候发生的巨大变化,这是极不可能的。使用生物类群的名称并不一定表明化石实际上代表了这些生物类群。
根据《国际藻类、真菌和植物命名法》,除非有充分的理由改变名称,否则首先使用的名称具有优先权。要进行这种更改,需要进行详细的研究,以积累适当的证据。从事这项工作的专家太少,因此古植物学文献中充斥着大量不恰当的植物部位名称,尤其是叶片,叶片自然会出现一系列变异,这些变异更多地与环境适应而非基因血统有关。花朵、花粉和种子在结构上往往更加稳定,不受环境影响,因此是遗传关系的更好指标,但在化石材料中,这些部分很少附着在叶片或母体植物的任何其他无性部分上。
本目录的目的是将北极植物化石材料的图像放到公共领域。它并不是要对北极白垩纪和古近纪植物命名法或分类法进行全面修订。
因此,所使用的名称应仅被视为在保存限制范围内表示特定形态的标签。由于各个化石的保存情况各不相同,而且很少完整,因此在使用这些名称时存在不同程度的不确定性。旧文献中使用的许多名称与活植物的名称直接相关,最常见的是属一级的名称(如 Alnus、Tilia、Menispermum、Quercus、Ziziphus 等)。这些名称不应被视为表明在白垩纪或古新世就存在与现在相同的植物。不过,由于我们有意不对命名法进行修订,因此这些旧名称与化石遗存特有的更合适的名称(如 Alnites、Tiliaephyllum、Menispermites、Quercophyllum、Ziziphoides 等)一起出现在本目录中。
4.1 样本标识符
美国地质调查局的标本被分配了标识符,标识符由地点编号、句号(.)和标本编号组成。 在对脉纹和边缘特征进行严格比较后,标本被归类为或多或少具有一系列一致特征的形态。 这些特征在形态描述中进行了总结。 如果这些形态与已公布的 "物种 "十分相似,则会参考已公布的名称,但在所有情况下,这些形态都以其独特的编号来识别。 现生(活体)植物的通用名称现在被认为不适合化石遗迹,因为化石遗迹缺少花或种子等诊断部分。但过去并非如此。如果化石标本使用了现生属名,有时会用单引号括起来。
4.2 正式名称
如果标本已被鉴定为以前描述过的形式,则使用正式的林奈双名。林奈双名(Linnaean binomial),正如 "双名 "一词所暗示的,由两部分组成:一是属名 (如水杉),二是与属名一起构成该物种特有的名称(如水杉属)。这第二个名称称为种的外标。有时不需要外标名,例如当我们想指某一属中的所有物种时,就只给出属名。如果我们可以将化石确定为某一属的化石,但不能确定为该属中的某一物种,我们就会使用属名后面的 “sp.”。当文中首次使用林奈二名或该二名是正式描述的一部分时,则会在该二名之后加上权威机构(首次描述或修订描述的作者姓名)。
在本目录中,正式二名仅用于那些我们可以确定为以前发表的种或属的标本。正式名称必须根据《国际藻类、真菌和植物命名规范》的规定在公认的科学出版物上发表过,才具有权威性。本图鉴不适合建立新的正式名称,因此在正式描述之前,我们的许多标本都被冠以临时或非正式名称。我们的非正式名称采用与林奈设计的完全不同的系统,通常由字母和数字组成形态描述符。
4.3 非正式名称
对于阿拉斯加的标本,当归属于已知的属和种有问题时,通常会使用形态型 “名称”。它们是非正式但有用的方法,或指代叶片类型,并基于一个能立即传达叶片形态的系统。
4.3.1 形态标识符
独特的数字形式标识符前缀有字母字符,大致表示叶片结构的主要特征。 希基的叶片分类方案)中的许多特征甚至在白垩纪叶片中也有关联。 例如,许多叶片具有全缘、凸脉和次脉;所有这些特征都与木兰科现存成员有关。 因此,白垩纪的被子植物形态被划分为人工群组,这些群组表现出同样一致的特征关联。 用于识别这些类群的代码来源于它们所表现出的建筑特征,在某些情况下也来源于现代亚类名称。 使用现代亚类名称并不是因为任何假定的系统发育关系,而仅仅是因为熟悉现存叶片形态就很容易理解其一般特征组合。
| 前缀 | 描述 |
|---|---|
| F | Ferns |
| E | Equisetites |
| GG | Gymnosperm - Ginkgophyte |
| GCO | Gymnosperm - Conifer |
| GCY | Gymnosperm - Cycadophyte |
| GU | Gymnosperm of unknown affinities |
| MA | Magnoliidae-like: camptodromous entire margined with intersecondary veins but lacking any pectinal veins |
| MAPE | Magnoliidae-like: as above but with pectinal veins, usually acrodromous |
| RAPE | Peltate or cordate leaves with numerous radiating pectinal veins |
| HATR | Trochodendroidean hamamelididaean-like cordate to ovate leaves usually with crenate (but maybe finely serrate to entire) margins, acrodromous pectinal veins |
| HAPLP | Hamamelididaean-like leaves with Platanoid teeth and peltate bases |
| HAPLTL | Hamamelididaean-like leaves with Platanoid teeth and truncate to lobate bases |
| HAPLD | Hamamelididaean-like leaves with Platanoid teeth and decureent bases |
| HASPTL | Hamamelididaean-like leaves with Spinose teeth and truncate or lobate bases |
| HASPD | Hamamelididaean-like leaves with Spinose teeth and decurrent bases |
| RO | Rosidaean-like leaves |
| DI | Dilleniidaean-like leaves |
| DIPE | Dilleniidaean-like leaves with pectinal veins |
部分或全部前缀的括号表示不确定性。U 表示亲缘关系不明。
如果在任何时候认为有必要更改某个形式的所属类别,只需更改前缀,同时保留唯一的形式编号即可。 只有当组成一个形式的标本全部分散到其他形式,而原来的形式被放弃时,才需要更改形式编号。
斯迈利收藏馆采用的是另一种系统,即给标本贴上非正式标签,如 "针叶树 18 "或 “圆锥花序 4”。这些名称是斯迈利在创建他的生物地层计划时所命名的。为了保持历史的连续性,我们保留了这些名称。有时,斯迈利会用属名来代替临时名称。同样,我们也保留了这些名称。不过要注意的是,有时斯迈利的属名与我们在直接获取俄罗斯标本比较后所指定的属名有所不同。
4.4 叶片描述术语
林奈(1753 年)将活体植物的分类合理化,大致形成了现在的形式。 与之前的其他学者一样,林奈主要依靠生殖器官的形态特征来划分植物类别。 与无性生殖器官相比,被子植物的花在物种水平上表现出的表型(环境)差异很小,因此可以提供一套基本稳定、持续可靠的特征。 这主要是因为生殖形态通常与特定的授粉机制有关,为了使空间上分离的个体之间成功受精,无论当地环境如何,授粉机制都必须是一致和兼容的。
另一方面,无性生殖器官必须在其所处的各种条件下尽可能有效地发挥作用,而那些能适当改变其形态、解剖和/或化学结构的植物则具有竞争优势。 无性器官的多变性阻碍了它们在系统学中的应用,因此这些器官(尤其是叶)的分类潜力一直被忽视。
然而,古植物学家很少有机会研究化石花。 花通常只生产数量有限的花朵,花朵结构脆弱,不太可能成为化石,花朵通常发育成果实或种子,这些果实或种子会被动物当作食物,或者发芽从而被毁坏。 因此,这类生殖器官在化石记录中比较罕见,而附着在生有叶子的树枝上的生殖器官更是少之又少。
因此,古植物学家长期以来一直对叶片残骸的分类潜力感兴趣,并试图用简洁明了的语言对其进行描述。 Ettingshausen (1861 年)是第一个为叶片结构制定综合术语的人,尽管没有得到严格遵守,但包括 Lesquereux (1878 年)、Berry (1916 年)和 Hollick (1930 年、1936 年)在内的一些学者部分地使用了这一术语。 随后,其他对古植物学感兴趣(但并非总是如此)的学者提出利用叶片特征进行分类学或环境研究: Lam(1925 年)、Lee(1948 年)、Mouton(1966 年,1970 年)、Ferguson(1971 年)、Hickey(1973 年,1979 年)、Dilcher(1974 年)、Dolph(1975 年,1976a 年,1976b 年)、Madler(1975 年)、Melville(1976 年)、Hill(1980 年)。 大多数建议的叶片特征分析方案都是作为书面描述的辅助工具(例如希基,1973 年,1979 年),但也有一些作者修改了描述术语,用于数值分析(多尔夫,1976 年 a,1976 年 b),或设计了原始方案,用于计算机技术(希尔,1980 年)。
希基(Hickey,1973 年)的著作或许是其中一个被广泛接受的描述性术语,随后,迪尔彻(Dilcher,1974 年)在其著作中加入了叶脉特征,并重新出版。 希基(1979 年)随后扩充了他的原始版本,经过多年的发展,该版本已成为现在的《叶片结构手册》(Ellis 等人,2009 年)。
叶片结构手册》中描述的方案虽然部分是基于化石叶片的经验,但无法充分涵盖白垩纪中期和晚期叶片结构的相互升级。这种明显的形态整合(包括脉纹模式)似乎是网状演化的结果,甚至在确定哪些是主脉、哪些是次脉这一最基本的层面上,也破坏了识别脉序的尝试。这是很不幸的,因为《叶片结构手册》中的许多后续描述性术语都依赖于正确的早期脉序分类。
为了克服这一问题,斯派塞(1986b)提出了茎脉术语,在本研究中,凡是尚未应用已发表描述的地方,都采用了这一术语。
4.5 叶脉
白垩纪被子植物表现出的特征组合跨越了现存科甚至目通常公认的界限。与此同时,晚白垩世的被子植物叶片显示出高度的形态可塑性。
这两个因素在很大程度上造成了白垩纪被子植物花粉和叶片系统学的混乱状态。就叶片而言,另一个问题是对叶片结构的描述不一致,而且往往图示不足。这些叶片所表现出的结构变化的连续性,以及限制性术语所带来的约束,常常迫使相似的叶片被归入明显不同的类群。因此,详细进化研究所需的微妙关系可能被掩盖了。为了根据叶脉的分支模式而不是叶脉粗细等方面的细微变化,为叶脉设计一个更客观的术语,我们设计了栉状叶脉术语。
茎脉是指离开叶片中脉(或另一条茎脉)的侧脉(或多条侧脉),它在叶背(叶背指远离中脉)支持着最多的附属分支。下面的叶脉样式图显示,左侧为无环状(craspedodromous)pectinal abmedial 叶脉,右侧为环状(brochidodromous)abmedial 叶脉。图(a)和(b)分别显示了强栉状脉和弱栉状脉。 栉脉可能起源于基部或基部上部,除背脉外,还带有内侧(朝向中脉)分支。